Oxidação dos Ácidos Graxos II (As Exceções)

A descrição padrão da oxidação dos ácidos graxos (beta-oxidação) se refere aos ácidos graxos "padrão": de cadeia longa (12 a 20 carbonos), saturados (sem dupla ligação), que se faz basicamente na mitocôndria (pela conveniência de usar os produtos, as acetilCoA, ali mesmo, no ciclo de Krebs, na maioria dos casos).
Acontece que não nos alimentamos somente com ácidos graxos de cadeia longa e saturados!
Mais ou menos a metade dos ácidos graxos da dieta, por exemplo, são insaturados, com uma ou mais dupla ligações, do tipo cis. Nesse caso (como no início da oxidação dos ácidos graxos saturados uma dupla ligação trans é criada entre o segundo e o terceiro carbono, alfa e beta), haverá a necessidade de uma isomerização (ou redução da dupla ligação) no momento em que o complexo enzimático "encontrar" a primeira ligação cis para que a beta-oxidação siga normalmente.

No caso de ácidos graxos com número ímpar de carbonos (menos comum), a beta-oxidação ocorre normalmente até o último passo, quando sobrarão não duas acetilCoA, mas uma acetilCoA e uma propionilCoA (uma coenzima A "segurando" um ácido graxo com 3 carbonos). Essa propionilCoA será transformada inicialmente em metil-malonilCoA, e depois em succinilCoA, um dos reagentes do ciclo de Krebs (que depois pode se transformar em oxalacetato e em glicose, como uma das vias da gliconeogênese).

Ácidos graxos de cadeia média (presentes principalmente em laticínios) são mais solúveis que os de cadeia longa (menor "cauda" apolar). Portanto, não são "bons candidatos" a serem "guardados" no tecido adiposo. Também não fazem a "rota habitual" das gorduras: são absorvidos pelo sistema portal e, ao chegarem ao fígado, sofrem beta-oxidação à acetilCoA (vão para uso, não para estoque, não viram triglicerídeos).
Os peroxissomos são organelas que talvez devessem ter uma plaquinha dentro da célula: "Cuidado! Material perigoso!". São eles os responsáveis pela oxidação de ácidos graxos de cadeia muito longa, com 24 a 26 carbonos (peroxissomos podem ser entendidos como "lisossomos especializados em oxidação de gorduras"). Ao atingir tamanhos menores de moléculas, os peroxissomos transferem o "trabalho mole" (a oxidação restante do ácido graxo encurtado) para a mitocôndria (organela com a qual o peroxissomo tem sempre "íntimo contato").
Uma característica importante do peroxissomo (daí a necessidade da "plaquinha") é utilizar uma outra enzima no início da oxidação, justamente uma oxidase, que irá doar os elétrons provenientes da oxidação para um oxigênio molecular (O2), criando o peróxido de hidrogênio (H2O2), composto altamente reativo e precursor dos chamados radicais livres, necessário nas respostas inflamatórias orgânicas (bem como precursor do HOCl, ácido hipocloroso, usado pelas células fagocíticas de defesa) que o peroxissomo "sabe como lidar" (pela presença das catalases, que neutralizam o peróxido). É a chamada compartimentalização (ou seja, a separação, nesse caso por motivos de segurança da célula) da produção de radicais livres.

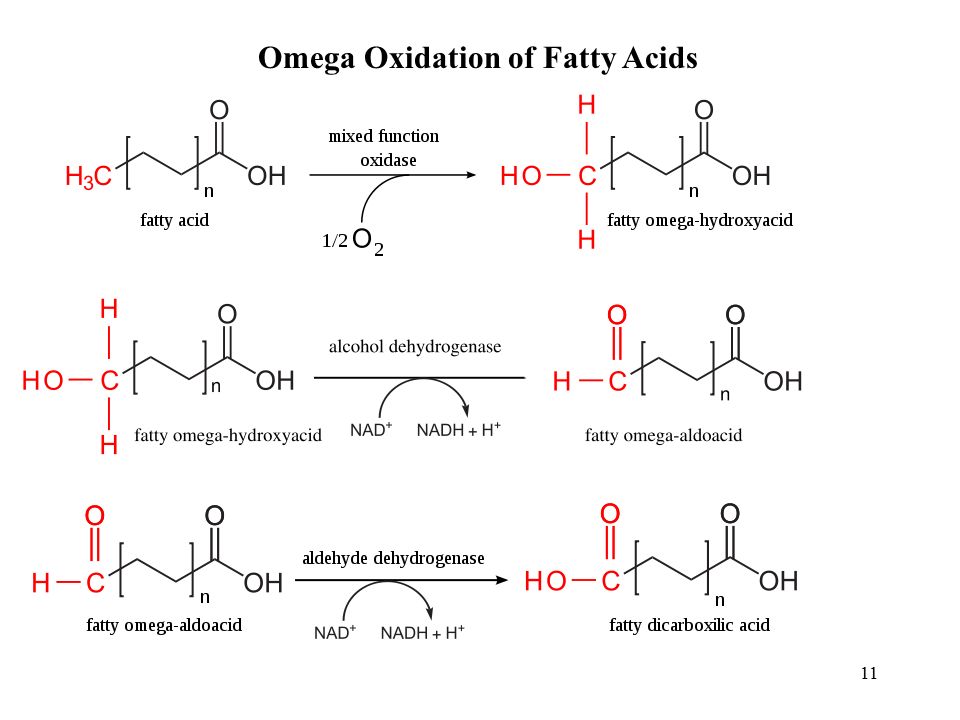
Ômega-oxidação: via alternativa de oxidação que, curiosamente, "corta" os carbonos pelo outro extremo da molécula do ácido graxo (o extremo ômega). Acontece no retículo endoplasmático, e é mais comum em situações de "defeito" na oxidação beta ou para oxidação de xenobióticos (moléculas estranhas ao organismo, mas semelhantes aos ácidos graxos).
A molécula é oxidada à álcool, depois à ácido dicarboxílico que, com suas duas "pontas carboxílicas" associadas à coenzima A, sofrem beta-oxidação em cada uma delas, restando no final dois ácidos carboxílicos encurtados (um deles, o succinato, entra no ciclo de Krebs - um passo além da succinilCoA, do final da oxidação dos ácidos graxos de cadeia ímpar, acima).
Comentários